А. ускорение световых и темновых реакций фотосинтеза
Б. использование световой энергии для синтеза органических веществ
В. расщепление органических веществ до неорганических
Г. участие в реакциях синтеза белка на рибосомах
Какой из перечисленных процессов происходит в световую фазу фотосинтеза?
А. образование глюкозы Б. синтез АТФ
В. поглощение CO 2 Г. все перечисленное
Назовите в хлоропласте участок, где происходят реакции темновой фазы фотосинтеза
А. наружная мембрана оболочки Б. вся внутренняя мембрана оболочки
В. граны Г. строма
30. Об условиях жизни древесных растений в разные годы можно узнать по толщине
А. Коры Б. Пробки
В. Лубяных волокон Г. Годичных колец
31. В пробирке с раствором хлорофилла фотосинтез не происходит, так как для этого процесса необходим набор ферментов, расположенных на
А. Кристах митохондрий Б. Гранах хлоропластов
В. Эндоплазматической сети Г. Плазматической мембране
Какие почки развиваются на листьях и корнях цветковых растений?
А. Придаточные Б. Верхушечные В. Пазушные Г. Боковые
33. Источником углерода, используемого растениями в процессе фотосинтеза, служит молекула
А. Угольной кислоты Б. Углеводорода
В. Полисахарида Г. Углекислого газа
Для улучшения дыхания корней культурных растений необходимо
А. Проводить прополку сорняков
Б. Систематически поливать растения
В. Периодически рыхлить почву вокруг растения
Г. Периодически подкармливать растения минеральными удобрениями
35. Приспособление растений к уменьшению испарения воды – наличие
А. Устьиц на верхней стороне листа
Б. Большого числа листовых пластинок
В. Широких листовых пластинок
Г. Воскового налета на листьях
36. Видоизмененный подземный побег многолетних растений с утолщенным стеблем, почками, придаточными корнями и чешуевидными листьями – это
А. Главный корень Б. Корневище
В. Боковой корень Г. Корнеклубень
Подземный побег отличается от корня наличием у него
А. Вегетативных почек
Б. Зоны проведения
В. Зоны всасывания
Г. корневых волосков
38. Какие удобрения усиливают рост зеленой массы растений?
А. Органические Б. Азотные
В. Калийные Г. Фосфорные
39. Свойство органов растений изгибаться под влиянием силы земного притяжения называют
А. Гидротропизмом Б. Фототропизмом
В. Геотропизмом Г. Хемотропизмом
40. Внешним сигналом, стимулирующим наступление листопада у растений, служит
А. Увеличение влажности среды
Б. Сокращение длины светового дня
В. Уменьшение влажности среды
Г. Повышение температуры среды
41. Затопление ранней весной полей пшеницы талыми водами иногда приводит к гибели всходов, так как при этом нарушается процесс
А. Фотосинтеза из-за недостатка кислорода
Б. Дыхания из-за недостатка кислорода
В. Поглощения воды из почвы
Г. Испарения воды
Часть В
В1(выберите несколько верных ответов из шести)
Значение транспирации
А. регулирует газовый состав внутри листа
Б. способствует передвижению воды
В. обеспечивает привлечение опылителей
Г. улучшает транспорт углеводов
Д. регулирует температуру листьев
Е. снижает удельный вес листвы
В2(выберите несколько верных ответов из шести)
Корневой чехлик выполняет функции
А. обеспечивает отрицательный геотропизм
Б. обеспечивает положительный геотропизм
В. облегчает проникновение корня в почву
Г. запасает питательные вещества
Д. защищает активно делящиеся клетки
Е. участвует в транспорте веществ
В3. Выберите несколько верных ответов
В чем состоит значение фотосинтеза?
А. в обеспечении всего живого органическими веществами
Б. в расщеплении биополимеров до мономеров
В. в окислении органических веществ до углекислого газа и воды
Г. в обеспечении всего живого энергией
Д. в обогащении атмосферы кислородом, необходимым для дыхания
Е. в обогащении почвы солями азота
В4. Установите соответствие между наиболее важными процессами и фазами фотосинтеза
В5. Установите правильную последовательность процессов фотосинтеза
А. возбуждение хлорофилла
Б. синтез глюкозы
В. соединение электронов с НАДФ + и Н +
Г. фиксация углекислого газа
Д. фотолиз воды
В6. Выберите несколько верных ответов
Выберите процессы, происходящие в световую фазу фотосинтеза
А. фотолиз воды Б. синтез углеводов
В. фиксация углекислого газа Г. синтез АТФ
Д. выделение кислорода Е. гидролиз АТФ
В7. Выберите несколько верных ответов
В темновую фазу фотосинтеза в отличие от световой происходит
А. фотолиз воды
Б. восстановление углекислого газа до глюкозы
В. синтез молекул АТФ за счет энергии солнечного света
Г. соединение водорода с переносчиком НАДФ +
Д. использование энергии молекул АТФ на синтез углеводов
Е. образование молекул крахмала из глюкозы
В8. Выберите несколько верных ответов
Органическое вещество состоит на 45% из углерода. Поэтому вопрос об источнике питания организмов углеродом чрезвычайно важен. Все организмы делят на автотрофные и гетеротрофные. Автотрофные организмы характеризуются способностью в качестве источника углерода использовать его минеральные формы, то есть синтезировать органическое вещество из неорганических соединений. Гетеротрофные организмы строят органическое вещество своего тела из уже имеющихся готовых органических соединений, то есть используют органические соединения как источник углерода. Для того, чтобы осуществить синтез органического вещества, необходима энергия. В зависимости от используемого соединения, а также от источников энергии, различают следующие основные типы питания углеродом и построения органических веществ.
Типы углеродного питания организмов
|
Тип питания |
Источник углерода |
Источник водорода |
Источник энергии |
|
I. Гетеротрофный |
Органическое вещество |
Органическое вещество |
Энергия окисления органических веществ |
|
П. Автотрофный 1. Фотосинтез |
Энергия света |
||
|
2.Бактериальный фотосинтез |
Н 2 S, Н 2 и др. |
Энергия света |
|
|
3. Хемосинтез |
Н 2 О, Н 2 S, Н 2 , |
Энергия окисления неорганических веществ |
Из всех перечисленных типов питания углеродом фотосинтез зеленых растений, при котором построение органических соединений идет за счет простых неорганических веществ (СО 2 и Н 2 О) с использованием энергии солнечного света, занимает совершенно особое место. Общее уравнение фотосинтеза:
6СО 2 + 12Н 2 О = С 6 Н 12 О 6 + 6О 2 + 6Н 2 О
Фотосинтез – это процесс, при котором энергия солнечного света превращается в химическую энергию. В самом общем виде это можно представить следующим образом: квант света (hv) поглощается хлорофиллом, молекула которого переходит в возбужденное состояние, при этом электрон переходит на более высокий энергетический уровень. В клетках фотоавтотрофов в процессе эволюции выработался механизм, при котором энергия электрона, возвращающегося на основной энергетический уровень, превращается в химическую энергию.
В процессе фотосинтеза из простых неорганических соединений (СО 2 , Н 2 О) строятся различные органические вещества. В результате происходит перестройка химических связей: вместо связей С–О и Н–О возникают связи С–С и С–Н, в которых электроны занимают более высокий энергетический уровень. Таким образом, богатые энергией органические вещества, которыми питаются и за счет которых получают энергию (в процессе дыхания) животные и человек, первоначально создаются в зеленом листе. Можно сказать, что практически вся живая материя на Земле является результатом фотосинтетической деятельности.
Почти весь кислород атмосферы фотосинтетического происхождения. Процессы дыхания и горения стали возможны только после того, как возник фотосинтез. Возникли аэробные организмы, способные усваивать кислород. На поверхности Земли процессы приняли биогеохимический характер, произошло окисление соединений железа, серы, марганца. Изменился состав атмосферы: содержание СО 2 и аммиака снизилось, а кислорода и азота возросло. Возникновение озонового экрана, который задерживает опасную для живых организмов ультрафиолетовую радиацию, также является следствием появления кислорода.
Для того, чтобы процесс фотосинтеза протекал нормально, к хлоропластам должен поступать СО 2 . Основным поставщиком служит атмосфера, где количество СО 2 составляет 0,03%. Для образования 1 г сахара необходимо 1,47 г СО 2 – такое количество содержится в 2500 л воздуха.
Углекислый газ поступает в лист растения через устьица. Некоторое количество СО 2 поступает непосредственно через кутикулу. При закрытых устьицах диффузия СО 2 в лист резко сокращается.
Наиболее примитивная организация фотосинтетического аппарата у зеленых бактерий и цианобактерий. У этих организмов функцию фотосинтеза выполняют внутрицитоплазматические мембраны или особые структуры – хлоросомы, фикобилисомы. У водорослей уже эволюционно возникли органеллы (хроматофоры), в которых сосредоточены пигменты, они разнообразны по форме (спиральные, лентовидные, пластинчатые, звездчатые). Высшие растения характеризуются вполне сформировавшимся типом пластид в форме диска или двояковыпуклой линзы. Приняв форму диска, хлоропласты становятся универсальным аппаратом фотосинтеза. Фотосинтез протекает в зеленых пластидах – хлоропластах. В лейкопластах синтезируется и отлагается в запасной крахмал, в хромопластах накапливаются каротиноиды.
Размер дисковидных хлоропластов высших растений колеблется от 4 до 10 мкм. Число хлоропластов обычно составляет от 20 до 100 на клетку. Химический состав хлоропластов достаточно сложен и может быть охарактеризован следующими средними данными (% на сухую массу): белок – 35-55; липиды – 20-30; углеводы – 10; РНК – 2-3; ДНК – до 0,5; хлорофилл – 9; каротиноиды – 4,5.
В хлоропластах сосредоточены ферменты, принимающие участие в процессе фотосинтеза (окислительно-восстановительные, синтетазы, гидролазы). В хлоропластах, так же как и в митохондриях, имеется своя белоксинтезирующая система. Многие из ферментов, локализованных в хлоропластах, являются двухкомпонентными. Во многих случаях простетическая группа ферментов – это различные витамины. В хлоропластах сосредоточены многие витамины и их производные (витамины группы В, К, Е, D). В хлоропластах находится 80% Fe, 70% Zn, около 50% Сu от всего количества этих элементов в листе.
Хлоропласты окружены двойной мембраной. Толщина каждой мембраны 7,5-10 нм, расстояние между ними 10-30 нм. Внутреннее пространство хлоропластов заполнено бесцветным содержимым – стромой и пронизано мембранами. Мембраны, соединенные друг с другом, образуют плоские замкнутые полости (пузырьки) – тилакоиды (греч. «тилакоидес» – мешковидный). В хлоропластах содержатся тилакоиды двух типов. Короткие тилакоиды собраны в пачки и расположены друг над другом, напоминая стопку монет. Эти стопки называются гранами, а составляющие их тилакоиды – тилакоидами гран. Между гранами параллельно друг другу располагаются длинные тилакоиды – тилакоиды стромы. Между отдельными тилакоидами в стопках гран имеются узкие щели. Тилакоидные мембраны содержат большое количество белков, участвующих в фотосинтезе. В составе интегральных мембранных белков имеется много гидрофобных аминокислот. Это создает безводную среду и делает мембраны стабильнее.
Для того, чтобы световая энергия могла быть использована в процессе фотосинтеза, необходимо ее поглощение фоторецепторами – пигментами. Фотосинтетические пигменты – это вещества, которые поглощают свет определенной длины волны. Не поглощенные участки солнечного спектра отражаются, что и обусловливает окраску пигментов. Так, зеленый пигмент хлорофилл поглощает красные и синие лучи, тогда как зеленые лучи, в основном, отражаются. Видимая часть солнечного спектра включает длины волн от 400 до 700 нм.
Состав пигментов зависит от систематического положения группы организмов. У фотосинтезирующих бактерий и водорослей пигментный состав разнообразен (хлорофиллы, бактериохлорофиллы, бактериородопсин, каротиноиды, фикобилины). Их набор и соотношение специфичны для различных групп организмов. Пигменты, сконцентрированные в пластидах, можно разделить на три группы: хлорофиллы, каротиноиды, фикобилины.
Важнейшую роль в процессе фотосинтеза играют зеленые пигменты –хлорофиллы. Французские ученые П.Ж. Пелетье и Ж. Кавенту (1818) выделили из листьев зеленое вещество и назвали его хлорофиллом (от греч. «хлорос» – зеленый и «филлон» – лист). В настоящее время известно около десяти хлорофиллов. Они отличаются по химическому строению, окраске, распространению среди групп организмов. У всех высших растений содержатся хлорофиллы a и b. Хлорофилл c обнаружен в диатомовых водорослях, хлорофилл d – в красных водорослях. Кроме того, известны бактериохлорофиллы (а, b, c, d), содержащиеся в клетках фотосинтезирующих бактерий. В клетках зеленых бактерий имеются бактериохлорофиллы с и d, в клетках пурпурных бактерий – бактериохлорофиллы a и b. Основными пигментами, без которых фотосинтез не идет, являются хлорофилл a для зеленых высших растений и водорослей, и бактериохлорофиллы – для бактерий.
Впервые точное представление о пигментах зеленого листа высших растений было получено благодаря работам крупнейшего российского ботаника М.С. Цвета (1872-1919). Он разработал новый хроматографический метод разделения веществ и выделил пигменты листа в чистом виде. Оказалось, что листья высших растений содержат хлорофилл a и хлорофилл b, а также каротиноиды (каротин, ксантофилл). Хлорофиллы, так же, как и каротиноиды, нерастворимы в воде, но хорошо растворимы в органических растворителях. Хлорофиллы a и b различаются по цвету: хлорофилл a имеет сине-зеленый оттенок, хлорофилл b – желто-зеленый. Содержание хлорофилла a в листе примерно в 3 раза больше по сравнению с хлорофиллом b. По химическому строению хлорофиллы – сложные эфиры дикарбоновой органической кислоты – хлорофиллина и двух остатков спиртов – фитола (С 20 Н 39 ОН) и метилового (СН 3 ОН). Эмпирическая формула хлорофилла С 55 Н 72 О 5 N 4 Мg (рис. 5.1 ).
Органическая дикарбоновая кислота хлорофиллин представляет собой азотсодержащее металлорганическое соединение, относящееся к магнийпорфиринам: (СООН) 2 = С 32 Н 30 ОN 4 Мg.
В хлорофилле водород карбоксильных групп замещен остатками двух спиртов – метилового СН 3 ОН и фитола С 20 Н 39 ОН, поэтому хлорофилл является сложным эфиром.
Рис. 5.1. Структурная формула хлорофилла а.
Хлорофилл b отличается тем, что содержит на два атома водорода меньше и на один атом кислорода больше (вместо группы СН 3 группа СНО). В связи с этим, молекулярная масса хлорофилла a – 893 и хлорофилла b – 907.
В центре молекулы хлорофилла расположен атом магния, который соединен с четырьмя атомами азота пиррольных группировок. В пиррольных группировках хлорофилла имеется система чередующихся двойных и простых связей. Это хромофорная группа хлорофилла, обусловливающая поглощение определенных лучей солнечного спектра и его окраску.
Еще К.А. Тимирязев обратил внимание на близость химического строения двух важнейших пигментов: зеленого – хлорофилла листьев и красного – гемина крови. Действительно, если хлорофилл относится к магнийпорфиринам, то гемин – к железопорфиринам. Сходство это служит еще одним доказательством единства всего органического мира.
Молекула хлорофилла полярна, ее порфириновое ядро обладает гидрофильными свойствами, а фитольный конец – гидрофобными. Это свойство молекулы хлорофилла обусловливает определенное расположение ее в мембранах хлоропластов. Порфириновая часть молекулы связана с белком, а фитольная цепь погружена в липидный слой.
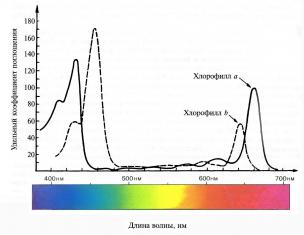
Хлорофилл способен к избирательному поглощению света. Спектр поглощения определяется его способностью поглощать свет определенной длины волны (определенного цвета). Для того чтобы получить спектр поглощения, К.А. Тимирязев пропускал луч света через раствор хлорофилла. Было показано, что хлорофилл в той же концентрации, как в листе, имеет две основные линии поглощения в красных и сине-фиолетовых лучах. При этом хлорофилл a в растворе имеет максимум поглощения 429 и 660 нм, тогда как хлорофилл b – при 453 и 642 нм (рис. 5.2).

Рис. 5.2. Спектры поглощения хлорофилла а и хлорофилла b
Наряду с зелеными пигментами в хлоропластах и хроматофорах содержатся пигменты, относящиеся к группе каротиноидов. Каротиноиды – это желтые и оранжевые пигменты алифатического строения, производные изопрена. Каротиноиды содержатся во всех высших растениях и у многих микроорганизмов. Это самые распространенные пигменты с разнообразными функциями. Каротиноиды, содержащие кислород, получили название ксантофиллы. Основными представителями каротиноидов у высших растений являются два пигмента – бета-каротин (оранжевый) С 40 Н 56 и ксантофилл (желтый) С 40 Н 56 О 2 . Каротин состоит из 8 изопреновых остатков. При разрыве углеродной цепочки пополам и образовании на конце спиртовой группы каротин превращается в 2 молекулы витамина А.
Бета-каротин имеет два максимума поглощения, соответствующие длинам волн 482 и 452 нм. В отличие от хлорофиллов каротиноиды не поглощают красные лучи, а также не обладают способностью к флуоресценции. Подобно хлорофиллу каротиноиды в хлоропластах и хроматофорах находятся в виде нерастворимых в воде комплексов с белками. Каротиноиды всегда присутствуют в хлоропластах, они принимают участие в процессе фотосинтеза. Поглощая световую энергию в определенных участках солнечного спектра, они передают энергию этих лучей на молекулы хлорофилла. Тем самым, они способствуют использованию лучей, которые хлорофиллом не поглощаются. Физиологическая роль каротиноидов не ограничивается их участием в передаче энергии на молекулы хлорофилла. Каротиноиды выполняют защитную функцию, предохраняя молекулы хлорофилла от разрушения на свету в процессе фотоокисления (рис. 5.3).

Рис. 5.3. Структурная формула бета-каротина
Фикобилины – красные и синие пигменты, содержащиеся у цианобактерий и красных водорослей. В основе химического строения фикобилинов лежат 4 пиррольные группировки. В отличие от хлорофилла у фикобилинов пиррольные группы расположены в виде открытой цепочки ( рис. 5.4).

Рис. 5.4. Структурная формула хромофорной группы фикоэритринов
Фикобилины представлены пигментами: фикоцианином, фикоэритрином и аллофикоцианином. Фикоэритрин – это окисленный фикоцианин. Красные водоросли, в основном, содержат фикоэритрин, а цианобактерии – фикоцианин. Фикобилины образуют прочные соединения с белками (фикобилинпротеиды). В отличие от хлорофиллов и каротиноидов, расположенных в мембранах, фикобилины концентрируются в особых гранулах (фикобилисомах), тесно связанных с мембранами тилакоидов. Фикобилины поглощают лучи в зеленой и желтой частях солнечного спектра. Это та часть спектра, которая находится между двумя основными линиями поглощения хлорофилла. Фикоэритрин поглощает лучи с длиной волны 495-565 нм, а фикоцианин – 550-615 нм. Сравнение спектров поглощения фикобилинов со спектральным составом света, в котором проходит фотосинтез у цианобактерий и красных водорослей, показывает, что они очень близки. Это позволяет считать, что фикобилины поглощают энергию света и, подобно каротиноидам, передают ее на молекулу хлорофилла, после чего она используется в процессе фотосинтеза. Наличие фикобилинов у водорослей является примером приспособления организмов в процессе эволюции к использованию участков солнечного спектра, которые проникают сквозь толщу морской воды (хроматическая адаптация).
Фотосинтез – это сложный многоступенчатый окислительно-восстановительный процесс, в котором происходит восстановление углекислого газа до углеводов и окисление воды до кислорода. В процессе фотосинтеза происходят не только реакции, идущие с использованием энергии света, но и темновые, не требующие непосредственного участия энергии света. Можно привести следующее доказательство существования темновых реакций в процессе фотосинтеза: фотосинтез ускоряется с повышением температуры. Отсюда прямо следует, что какие-то этапы этого процесса непосредственно не связаны с использованием энергии света. Процесс фотосинтеза включает следующие этапы: 1) фотофизический; 2) фотохимический (световой); 3) ферментативный (темновой).
Согласно законам фотохимии, при поглощении кванта света атомом или молекулой какого-либо вещества электрон переходит на другую, более удаленную орбиталь, то есть на более высокий энергетический уровень (рис. 5.5).

Рис. 5.5. Переходы между возбужденными состояниями хлорофилла после поглощения квантов синего и красного света
Наибольшей энергией обладает электрон, отдаленный от ядра атома и находящийся на достаточно большом расстоянии от него. Каждый электрон переходит на более высокий энергетический уровень под влиянием одного кванта света, если энергия этого кванта равна разнице между этими энергетическими уровнями. Все фотосинтезирующие организмы содержат какой-либо тип хлорофилла. В молекуле хлорофилла два уровня возбуждения. Именно с этим связано и то, что он имеет две основные линии поглощения. Первый уровень возбуждения обусловлен переходом на более высокий энергетический уровень электрона в системе сопряженных двойных связей, а второй – с возбуждением неспаренных электронов атомов азота и кислорода в порфириновом ядре. При поглощении света электроны переходят в колебательное движение и перемещаются на следующие орбитали с более высоким энергетическим уровнем.
Наиболее высокий энергетический уровень – это второй синглетный уровень. Электрон переходит на него под влиянием сине-фиолетовых лучей, кванты которых содержат больше энергии.
В первое возбужденное состояние электроны могут переходить, поглощая более мелкие кванты красного света. Время жизни на втором уровне составляет 10 -12 с. Это время настолько мало, что на его протяжении энергия электронного возбуждения не может быть использована. Через этот короткий промежуток времени электрон возвращается в первое синглетное состояние (без изменения направления спина). Переход из второго синглетного состояния в первое сопровождается некоторой потерей энергии (100 кДж) в виде теплоты. Время жизни в первом синглетном состоянии немного больше (10 -9 или 10 -8 с). Наибольшим временем жизни (10 -2 с) обладает триплетное состояние. Переход на триплетный уровень происходит с изменением спина электрона.
Из возбужденного, первого синглетного и триплетного состояния молекула хлорофилла также может переходить в основное. При этом ее дезактивация (потеря энергии) может проходить:
1) путем выделения энергии в виде света (флуоресценция и фосфоресценция) или в виде тепла;
2) путем переноса энергии на другую молекулу пигмента;
3) путем затрачивания энергии на фотохимические процессы (потеря электрона и присоединение его к акцептору).
В любом из указанных случаев молекула пигмента дезактивируется и переходит на основной энергетический уровень.
Хлорофилл имеет две функции – поглощение и передачу энергии. При этом основная часть молекул хлорофилла – более 90% всего хлорофилла хлоропластов входит в состав светособирающего комплекса (ССК). Светособирающий комплекс выполняет роль антенны, которая эффективно поглощает свет и переносит энергию возбуждения к реакционному центру. Кроме большого числа (до нескольких сотен) молекул хлорофилла ССК содержит каротиноиды, а у некоторых водорослей и цианобактерий – фикобилины, которые увеличивают эффективность усвоения света.
В процессе эволюции в растениях выработался механизм, позволяющий наиболее полно использовать кванты света, падающие на лист подобно каплям дождя. Механизм этот заключается в том, что энергия квантов света улавливается 200-400 молекулами хлорофилла и каротиноидами ССК и передается одной молекуле – реакционному центру. Расчеты показали, что в одном хлоропласте до 1 млрд молекул хлорофилла. Теневыносливые растения имеют, как правило, больший размер ССК по сравнению с растениями, растущими в условиях высокой освещенности. В реакционных центрах в результате фотохимических реакций образуются первичные восстановитель и окислитель. Они затем вызывают цепь последовательных окислительно-восстановительных реакций. В итоге энергия запасается в виде восстановленного никотинамидадениндинуклеотидфосфата (НАДФ Н+) и аденозинтрифосфата (АТФ), который синтезируется из аденозиндифосфата (АДФ) и неорганической фосфорной кислоты за счет реакции фотосинтетического фосфорилирования. Следовательно, НАДФ Н+ и АТФ – основные продукты световой фазы фотосинтеза. Таким образом, в первичных процессах фотосинтеза, связанных с поглощением молекулой хлорофилла кванта света, важную роль играют процессы передачи энергии. Фотофизический этап фотосинтеза заключается в том, что кванты света поглощаются и переводят молекулы пигментов в возбужденное состояние. Затем эта энергия переносится на реакционный центр, осуществляющий первичные фотохимические реакции: разделение зарядов. Дальнейшее превращение энергии света в химическую энергию проходит ряд этапов, начиная с окислительно-восстановительных превращений хлорофилла и включая как фотохимические (световые), так и энзиматические (темновые) реакции.
То есть фотосинтез включает преобразование энергии (явление, получившее название светового процесса) и превращение вещества (темновой процесс). Световой процесс происходит в тилакоидах, темновой – в строме хлоропластов. Два процесса фотосинтеза выражаются отдельными уравнениями:
12Н 2 О =12Н 2 + 6О 2 + энергия АТФ (световой процесс).
Из этого уравнения видно, что кислород, выделяемый при фотосинтезе, образуется при разложении молекул воды. Кроме того, световая энергия используется на синтез аденозинтрифосфорной кислоты (АТФ) в ходе фотофосфорилирования.
6СО 2 + 12Н 2 + энергия АТФ = С 6 Н 12 О 6 + Н 2 О (темновой процесс)
В темновых реакциях используются продукты, накопленные в световой фазе. Суть темновых реакций сводится к фиксации СО 2 и включению его в молекулу сахара. Этот процесс получил название цикла Кальвина по имени американского биохимика, подробно изучившего последовательность темновых реакций. Использование воды в качестве источника водорода для синтеза органических молекул дало растениям в процессе эволюции большое преимущество в силу повсеместного ее присутствия (вода является самым распространенным минералом на Земле).
Поскольку весь кислород фотосинтеза выделяется из воды, итоговое уравнение принимает вид:
6СО 2 + 12Н 2 О + hv = С 6 Н 12 О 6 + 6О 2 + 6Н 2 О
Вода в правой части уравнения не подлежит сокращению, так как ее кислород имеет иное происхождение (из СО 2). Следовательно, фотосинтез – это окислительно-восстановительный процесс, в котором вода окисляется до молекуляргого кислорода (О 2) , а углекислый газ восстанавливается водородом воды до углеводов.
По завершении каждого цикла образуется конечный продукт: одна молекула сахара, который ложится в основу первичного органического вещества, образующегося при фотосинтезе.
Содержание статьи
ЦИКЛ УГЛЕРОДА, круговорот углерода, – циклическое перемещение углерода между миром живых существ и неорганическим миром атмосферы, морей, пресных вод, почвы и скал. Это один из важнейших биогеохимических циклов, включающий множество сложных реакций, в ходе которых углерод переходит из воздуха и водной среды в ткани растений и животных, а затем возвращается в атмосферу, воду и почву, становясь снова доступным для использования организмами. Поскольку углерод необходим для поддержания любой формы жизни, всякое вмешательство в круговорот этого элемента влияет на количество и разнообразие живых организмов, способных существовать на Земле.
Источники и резервы углерода.
Основной источник углерода для живых организмов – это атмосфера Земли, где данный элемент присутствует в виде диоксида углерода (углекислого газа, СО 2). В течение многих миллионов лет концентрация СО 2 в атмосфере, по-видимому, существенно не менялась, составляя ок. 0,03% веса сухого воздуха на уровне моря. Хотя доля СО 2 невелика, его абсолютное количество поистине огромно – ок. 750 млрд. т. В атмосфере СО 2 переносится ветрами как в вертикальном, так и в горизонтальном направлениях.
Диоксид углерода присутствует в воде, где он легко растворяется, образуя слабую угольную кислоту Н 2 СО 3 . Эта кислота вступает в реакции с кальцием и другими элементами, образуя минералы, называемые карбонатами. Карбонатные породы, например известняк, находятся в равновесии с диоксидом углерода, который содержится в контактирующей с ними воде. Аналогичным образом количество СО 2 , растворенного в океанах и пресных водах, определяется его концентрацией в атмосфере. Общее количество растворенных и осадочных углеродсодержащих веществ оценивается примерно в 1,8 трлн. т.
Углерод в соединении с водородом и другими элементами является одним из основных компонентов клеток растений и животных. Например, в организме человека он составляет ок. 18% массы тела. Многочисленность и очень широкое распространение живых организмов не позволяют удовлетворительно оценить общее содержание в них углерода. Можно, однако, приблизительно оценить суммарное количество углерода, связываемого растениями, а также выделяемого в процессе дыхания растений, животных и микроорганизмов. Установлено, что зеленые растения поглощают в год ок. 220 млрд. т CO 2 . Почти такое же количество этого вещества выделяется в неорганическую среду в процессе дыхания всех живых организмов, а также в результате разложения и сгорания органических веществ.
При определенных условиях разложения и сгорания созданных живыми организмами веществ не происходит, что ведет к накоплению углеродсодержащих соединений. Так, например, древесина живых деревьев может быть на 3–4 тысячелетия надежно защищена от микробного разложения и от пожара корой, способной противостоять действию микробов и огня. Древесина же, попавшая в торфяное болото, сохраняется еще дольше. В обоих случаях связанный в ней углерод оказывается как бы в ловушке и надолго выводится из круговорота. В условиях, когда органическое вещество оказывается захороненным и изолированным от воздействия воздуха, оно разлагается только частично и содержащийся в нем углерод сохраняется. Если впоследствии в течение миллионов лет эти органические остатки подвергаются давлению вышележащих отложений и нагреванию за счет земного тепла, значительная часть его превращается в ископаемое топливо, например в каменный уголь или нефть. Ископаемое топливо образует природный резерв углерода. Несмотря на интенсивное его сжигание, начавшееся с 1700-х годов, неизрасходованными еще остаются примерно 4,5 трлн. т.
Фотосинтез.
Основной путь, посредством которого углерод из мира неорганического перемещается в мир живого, – это осуществляемый зелеными растениями фотосинтез. Данный процесс представляет собой цепь реакций, в ходе которых растения поглощают из атмосферы или воды диоксид углерода, связывая его молекулы с молекулами специального вещества – акцептора СО 2 . В ходе других реакций, идущих с потреблением солнечной (световой) энергии, происходит расщепление молекул воды и использование высвобождающихся ионов водорода и связанного СО 2 в синтезе богатых углеродом органических веществ, в том числе акцептора СО 2 .
На каждую молекулу СО 2 , которую поглощает растение, чтобы синтезировать органические вещества, выделяется молекула кислорода, образованная при расщеплении воды. Предполагается, что именно таким путем образовался весь свободный кислород атмосферы. Если бы процесс фотосинтеза на Земле внезапно прекратился и нарушился углеродный цикл, то, согласно имеющимся расчетам, весь свободный кислород исчез бы из атмосферы примерно за 2000 лет.
Другие реакции.
Зеленое растение использует углерод образуемых им органических веществ разными способами. Например, он может накапливаться в составе крахмала, запасаемого в клетках, или целлюлозы – основного структурного материала растений и питательного вещества для многих других организмов. И крахмал и целлюлоза усваиваются в качестве пищи только после расщепления на составляющие их 6-углеродные сахара (т.е. сахара, содержащие по шесть атомов углерода в молекуле). В отличие от крахмала – нерастворимого высокомолекулярного соединения – 6-углеродные сахара легко растворимы и, перемещаясь по растению, служат источником энергии и материалом для роста и обновления клеток, а также для их восстановления в случае повреждений. Проростки, например, расщепляют запасенные в семени крахмал и жиры, получая из них более простые органические вещества, используемые в процессе клеточного дыхания (для высвобождения их энергии) и для роста.
У животных поглощенная пища подвергается аналогичному процессу переваривания. Прежде чем ее основные компоненты могут быть усвоены, они должны быть преобразованы: углеводы – в 6-углеродные сахара, жиры – в глицерин и жирные кислоты, белки – в аминокислоты. Эти продукты переваривания служат животному источниками энергии, высвобождаемой при дыхании, а также строительными блоками, необходимыми для роста организма и обновления его компонентов. Подобно растениям, животные способны переводить питательные вещества в форму, удобную для запасания. Аналог крахмала у животных – это гликоген, образуемый из излишков 6-углеродных сахаров и накапливаемый в качестве энергетического резерва в печени и мышечных клетках. Избыток сахара может превращаться также в жирные кислоты и глицерин, которые вместе с такими же веществами, поступающими с пищей, используются для синтеза жиров, накапливаемых в ткани. Таким образом, процессы синтеза обеспечивают запасание богатых углеродом и связанной энергией веществ, что позволяет организму выживать в периоды нехватки пищи.
После своей смерти растения и животные становятся пищей для т.н. редуцентов – организмов, осуществляющих разложение органического вещества. Большая часть редуцентов представлена бактериями и грибами, клетки которых выделяют наружу, в свое непосредственное окружение, небольшие количества пищеварительной жидкости, расщепляющей субстрат, а затем потребляют продукты такого «переваривания». Как правило, редуценты имеют ограниченный набор ферментов и соответственно используют в качестве пищи и источника энергии только немногие типы органических веществ. Обычные дрожжи, например, перерабатывают только 6- и 12-углеродные сахара, содержащиеся в разрушенных клетках перезрелых фруктов или в густом (с мякотью) соке, полученном при их раздавливании. Однако при достаточной длительности воздействия разнообразных редуцентов все углеродсодержащие вещества растений или животных в конце концов разрушаются до диоксида углерода и воды, а высвобожденная энергия используется организмами, осуществляющими разложение. Многие искусственно синтезированные органические соединения тоже подвержены биологическому разрушению (биодеградации) – процессу, в ходе которого редуценты получают энергию и необходимый строительный материал, а в атмосферу выделяется углерод в форме диоксида углерода.
§ 20. Ассимиляция углерода зелеными растениями
Химический состав разных растений неодинаков, но в среднем можно принять, что на долю углерода в нем приходится около 45, кислорода - 42, водорода - 6,5, азота - 1,5% и золы около 5 %. Таким образом, если не считать золы, тело растения, в том числе и дерева, состоит в основном из четырех химических элементов: углерода, кислорода, водорода и азота. Откуда же берутся эти элементы? Кислород и водород растение получает из воды. Об источнике азота будет сказано несколько позже. Откуда же берет дерево углерод, составляющий почти половину его сухого веса?
В 1771 г. английский ученый Д. Пристлей проделал свой знаменитый опыт с мышью и веткой зеленого растения. Мышь погибала под стеклянным колпаком. Если, же вместе с нею под колпак помещали ветку зеленого растения (мяты), она оставалась живой. Д. Пристлей сделал вывод, что зеленые растения «исправляют» воздух, испорченный дыханием животных. Однако скоро появились опровержения этого открытия, так как у других ученых получалось, что зеленые растения, как и животные, не исправляют воздух, а делают его негодным для дыхания и горения. Возникшее противоречие разрешил в 1779 г. Ян Ингенгуз, установивший, что зеленые растения действительно очищают воздух, но только на свету, где зеленые растения, очищая воздух, поглощают углекислый газ (в этом состоит их воздушное питание) и выделяют кислород. Объемы поглощаемого СО2 и выделяемого О2 равны. Одновременно с углекислым газом растение использует еще и воду. Этот процесс был назван фотосинтезом (от греческих слов фотос - свет и синтезис - образование сложных химических соединений из простых), или ассимиляцией углекислоты. Суммарная реакция фотосинтеза такова:
6 CO 2 + 6Н2 О → свет С6Н1206 + 602
Как видим, из 6 молекул углекислого газа и 6 молекул воды с поглощением солнечной энергии получается 1 молекула углевода и выделяются в атмосферу 6 молекул свободного кислорода. Однако в приведенной реакции даны лишь начало и конец процесса. На самом же деле он проходит значительно сложнее, имеет множество промежуточных этапов, изучение которых продолжается и сейчас. Эта реакция имеет неоценимое значение, так как Жизнь на Земле возможна лишь благодаря фотосинтезу.
При создании каждой молекулы сахара в ней оказывается, законсервированной солнечная энергия, перехваченная зеленым листом. В целом, по подсчетам X. Райнботе , ежегодно растения запасают в процессе фотосинтеза в виде углеводов столько энергии, сколько расходуют ее 100 тыс. больших городов в течение 100 лет.
Рис. 53. Определение фотосинтеза при помощи стеклянной колбы - : 1 - колба; 2 - термометр; 3 - пробка
Все запасы на Земле каменного угля, торфа, нефти, горючих сланцев представляют собой законсервированную солнечную энергию, полученную в процессе фотосинтеза растений, живших на Земле очень давно. Великий русский ученый зев отметил, что зеленое растение - промежуточное звено между животным (в том числе и человеком) и Солнцем, и его роль действительно является космической. Он писал, что пища служит источником силы в нашем организме потому только, что она не что иное, как консервы солнечных лучей.
Как же можно обнаружить, а еще лучше измерить фотосинтез дерева или хотя бы его части - отдельной ветки, листа? Очень часто учитывают количество СО2, поглощенное растением из воздуха, где содержание углекислого газа составляет всего лишь 0,03 % по объему, или 3 части на 10 тыс. частей воздуха.
Стеклянную колбу (способ предложен и; емкостью около 3 л надевают на ветку дерева при помощи разрезной пробки, герметически закрывающей входное отверстие (рис. 53). Колбу оставляют на 20 мин, не более, иначе в ней будет поглощена веткой вся углекислота и фотосинтез остановится. После этого колбу быстро снимают с ветки и тут же в нее наливают определенное количество едкой щелочи, обычно так называемой баритовой воды - Ba(OH)2. Щелочь жадно поглощает всю оставшуюся в колбе CO2, которую не успело усвоить растение. Для определения этой величины щелочь из колбы титруют, т. е. приливают к ней по каплям щавелевую кислоту до наступления между кислотой и щелочью химического равновесия. Если пустую колбу выдержать несколько минут на воздухе рядом с опытной веткой, а затем проделать с ней то же, что и с первой колбой, то это покажет общее количество СО2 в колбе в начале опыта. Отняв от этой величины первую, получают количество СО2, поглощенное веткой дерева. Далее определяют площадь листьев опытной ветки. Интенсивность фотосинтеаа выражают в мг СО2, усвоенных 1 дм2 поверхности листьев в течение 1 ч (мг./дм2 ч).
У хвойных, особенно, например, у ели, площадь хвоинок определил трудно, тогда берут вес хвои (сухой или сырой) и выражают фотосинтез в мг С02 на 1 г хвои в 1 ч.
Основные органы фотосинтеза - листья (хвоя). Фотосинтез идет и в молодых, еще зеленых побегах, содержащих хлоропласты, но слабее, чем в листьях. Как уже отмечалось, фотосинтез происходит в зеленых пластидах - хлоропластах, имеющих очень сложное строение. В хлоропластах содержится зеленый пигмент хлорофилл, заключенный в особые частицы - граны, погруженные в бесцветную основу хлоропласта - строму, состоящую из белков. При погружении зеленых листьев в спирт, бензин или эфир хлорофилл легко переходит в раствор, который при этом зеленеет, а листья обесцвечиваются. Перейдя из листа в спирт, хлорофилл теряет способность к фотосинтезу, который происходит лишь при тесном взаимодействии хлорофилла с белковыми веществами стромы хлоропласта. Кроме того, даже при малейшем нарушении структуры хлоропласта (например, если слегка прокатить по зеленому листу растения стеклянной палочкой) фотосинтез также прекращается. Все это показывает, насколько сложно устроен хлоропласт.
Хлорофилл состоит из сложного эфира дикарбоновой кислоты и 2 спиртов - метилового и высокомолекулярного непредельного спирта фитола. Этот зеленый - пигмент представлен в листе двумя разновидностями: хлорофиллом «a» (C55Н72O5N4Mg) и хлорофиллом «б» (C55Н70O6N4Mg). Кроме того, в листьях растений имеется еще оранжевый пигмент каротин (С40Н56), содержащийся также в изобилии в оранжево-красных корнеплодах моркови (латинское название которой даукус карота и дало название пигменту), и желтый пигмент ксантофилл (С40Н56O2).
Главную роль в процессе фотосинтеза играет хлорофилл. Он поглощает солнечные лучи и направляет их энергию на восстановление СО2 до углеводов, причем различные лучи солнечного света поглощаются хлорофиллом неодинаково. Это хорошо видно, если пропустить луч света через раствор хлорофилла в спирте, а затем через стеклянную призму спектроскопа. Солнечный свет состоит из семи лучей, видимых нашим глазом (красных, оранжевых, желтых, зеленых, голубых, синих и фиолетовых). Общая картина поглощения различных лучей хлорофиллом называется его спектром. Наиболее интенсивно хлорофилл поглощает часть красных лучей с длиной волны 650-680 нанометров (нанометр - одна миллиардная доля метра) и сине-фиолетовые с длиной волны около 470 нанометров. Зеленые лучи почти полностью отражаются хлорофиллом, что и придает ему изумрудно-зеленую окраску.
Для образования хлорофилла нужны определенные условия. Одно из них - свет. Клубни картофеля, хранящегося в темном подвале, образуют слабые, бледные, сильно вытянутые побеги. Такие растения называют этиолированными. При выставлении на свет они быстро зеленеют. Следующим необходимым условием для образования хлорофилла является наличие в почве солей железа. На щелочных почвах эти соли переходят в нерастворимую форму и становятся недоступными для корней растения. Хотя они при этом и не вытягиваются, но выглядят такими же бледными, как и при этиоляции. Такое явление получило название хлороза. На основании этого долгое время считали, что железо входит в состав хлорофилла. Как мы теперь знаем, это не так, но железо принимает активное участие в синтезе хлорофилла как катализатор, хотя количество его ничтожно. Так, если на бледном листе хлоротического растения нанести царапину гвоздем, то она сразу же позеленеет, остальные же участки листа останутся такими же бледными, как и были. Поздней холодной весной можно наблюдать желтую, долго не зеленеющую молодую траву. Образование хлорофилла в ней задерживается низкой температурой.
Солнечный свет, попадая на молекулу хлорофилла, приводит ее в возбужденное состояние, в результате чего образуется аденозитрифосфорная кислота (АТФ)-универсальный источник энергии в клетке. впервые доказал, что фотосинтез наиболее интенсивно идет в лучах, наиболее активно поглощаемых хлорофиллом, т. е. в красных и сине-фиолетовых.
Лист растения по внешнему строению и внутренней структуре является органом, специально приспособленным для фотосинтеза. Он имеет вид тонкой пластинки, хорошо улавливающей свет, состоит из мякоти, пронизанной густой сетью жилок. С поверхности листа легко сдирается кожица, покрывающая, как отмечалось, лист одним слоем клеток и несущая на себе жировую пленку - кутикулу. Густо разветвленная сеть жилок хорошо снабжает каждый участок листа водой, поступающей сюда из почвы по черешку, стволу и корням. По жилкам же оттекают в ствол образующиеся в листе сахара. Эти два встречных потока передвигаются по разным частям жилки (ксилеме и флоэме), не мешая друг другу. Кожица листа совершенно прозрачна и через нее в лист легко проходит свет.
Как уже отмечалось, одним из конечных продуктов фотосинтеза является сахар - глюкоза, другим - кислород. Сахар, накопившийся в течение дня, ночью оттекает из листа. Днем тоже происходит отток, но более слабый, и обычно часть образовавшегося сахара, чтобы не мешать дальнейшему процессу фотосинтеза, здесь же, в листе, выводится из реакции, превращаясь в крахмал. Это первый видимый продукт фотосинтеза, который легко обнаружить в листе действием йода, от которого крахмал синеет.
Взрослое дерево в почве ветвится многочисленными корнями, следуя за влагой и растворенными в ней питательными минеральными солями. Верхняя часть дерева - крона, несущая листья или хвою, устремляется навстречу солнечному свету. Листья на дереве располагаются в виде так называемой листовой мозаики. Это значит, что каждый лист не затеняет соседние или находящиеся непосредственно под ним.
Казалось бы, раз растения так стремятся за светом, то наилучшего своего развития они должны достичь при максимальном освещении. На самом деле это не так, все обстоит гораздо сложнее. Для фотосинтеза и общего развития разным растениям требуется различное количество солнечного света. Особенно велико это различие у травянистых растений. Например, известный степной сорняк подсвекольник, или щирица, может расти только на совершенно открытых местах и даже малейшее затенение заметно подавляет его развитие. Такие растения называются светолюбивыми. В то же время часто встречающийся в лиственных лесах дубровник не переносит открытых мест и наилучшего своего развития достигает при значительном затенении, когда щирица уже почти отмирает от недостатка света. Такие виды, как дубровник, называют тенелюбивыми. Конечно, эти два растения представляют собой две противоположности по отношению к свету. Между ними находятся виды, занимающие промежуточное положение, способные переносить большую или меньшую степень затенения. Такие растения называются теневыносливыми.
Если говорить только о древесных растениях, то среди них нет крайних тенелюбов, вроде дубровника, и их можно разбить на две категории: светолюбивые и теневыносливые. Однако есть очень светолюбивые деревья и крайне теневыносливые, а между ними - промежуточные виды. Все они отличаются друг от друга мерой (степенью) свотолюбия или теневыносливости. Для определения степени светолюбия немецкий ученый И. Визнер поступал следующим образом. Он находил наиболее затененный участок, где еще встречался данный вид растения, измерял освещенность в полдень и сравнивал ее с освещенностью на совершенно открытом месте, принятой за единицу. Получалась дробь, показывающая, какой частью полного солнечного освещения может довольствоваться данный вид: чем меньше дробь, тем более теневыносливым является растение, и наоборот. По И. Визнеру, наши наиболее распространенные древесные породы могут существовать при следующих степенях освещенности (в долях от полного солнечного света):
Лиственница..1/5
Берёза……….1/7
Сосна……......1/9
Дуб…………..1/25
Ель……………1/28
Клен………….1/55
Бук……………1/60
Самшит………..1/100
Полученный ряд представляет собой шкалу светолюбия группы древесных пород, в начале которой стоят самые светолюбивые, в конце - наиболее теневыносливые. Эта шкала может быть и более полной, включающей большее число видов, но принцип построения ее остается тем же. В приведенной шкале самшит оказался в 20 раз теневыносливее лиственницы, а лиственница соответственно в 20 раз светолюбивее самшита. Но это еще не предел теневыносливости зеленых растений. Так, водоросли , живущие в почве на глубине до 10 см, могут довольствоваться 1/2500 частью света.
Для измерения интенсивности света И. Визнер пользовался фотографической бумагой, которую выставлял на свет и по скорости ее потемнения (замеченной по секундомеру) судил об относительной освещенности данного места. Если, к примеру, на открытой поляне бумага темнела в течение 1 мин, а под пологом соседнего леса для этого требовалось 2 мин, то можно было принять освещенность под пологом равной половине освещенности на открытом месте. Сейчас освещенность измеряется более точно специальными приборами (люксметрами и др.).
Светолюбивые и теневыносливые деревья отличаются внешне. Светолюбивые виды имеют резкую крону, стволы их быстро очищаются от нижних сучьев, которые отмирают вследствие светового голодания. Такова лиственница, крона которой со всех сторон так пронизана светом, что он достигает каждой отдельной хвоинки. Белоствольная береза, кажется, сама излучает свет, отражая его стволом и ветвями. Крона ее также редкая, и свет проникает ко всем листьям. Под пологом лиственничного и березового леса остается еще достаточно света для образования густого травянистого напочвенного покрова.
Теневыносливые породы имеют густые пирамидальные кроны с нижними ветвями, спускающимися почти до поверхности почвы. Часто в таких насаждениях почти нет травяного напочвенного покрова. Под пологом леса из таких пород царит полумрак. Таковы еловые, пихтовые и буковые леса.
Интенсивность света неодинакова даже в различных частях одного и того же дерева. С наружной стороны кроны света больше, чем с внутренней. Поэтому и листья снаружи и внутри кроны существенно отличаются по анатомическому строению. В первом случае развивается так называемый световой тип листа, во втором - теневой. Главное отличие этих листьев состоит в том что в световом листе сильнее развита столбчатая ткань, лучше приспособленная для использования прямых солнечных лучей. Она располагается в 2, иногда даже и в 3 слоя. В теневом листе столбчатой ткани всего 1 ряд (нередко она отсутствует), зато гораздо мощнее губчатая ткань (см. рис. 47), полнее улавливающая рассеянный свет, господствующий внутри кроны. Этому же способствуют большое количество хлорофилла и более крупные хлоропласты в листьях теневыносливых пород (бук, тис), по сравнению со светолюбивыми (лиственница, акация белая).
Как же идет фотосинтез у светолюбивых и теневыносливых пород при различной степени освещенности? Это очень. наглядно показал (см. табл.). Из данных таблицы видно, что у лиственных пород фотосинтез идет интенсивнее, чем у хвойных, при всех степенях освещения, хотя из хвойных лиственница ассимилирует почти так же, как лиственные, а из лиственных дуб так же слабо, как и хвойные. Более высокий фотосинтез у лиственных связан со структурой плоского листа, лучше приспособленного к ассимиляций (плоская пластинка, лучше улавливающая свет, хорошо развитая столбчатая ткань и др.). Среди хвойных наиболее интенсивно ассимилирует пихта, ель отличается слабым фотосинтезом при любой интенсивности света. Из лиственных наибольший фотосинтез у липы и наименьший у дуба. Таблица показывает также, что теневыносливые породы наиболее заметно отличаются от светолюбивых при слабом освещении, когда первые ассимилируют энергичнее. При дальнейшем увеличении света (до 30 %) .различия в фотосинтезе у светолюбивых и теневыносливых пород уменьшаются. Наконец, при приближении к полному солнечному освещению фотосинтез у светолюбивых пород (особенно хвойных) становится выше, чем у теневыносливых.
Влияние степени освещенности на интенсивность фотосинтеза (мгС09 на 1 г сырого веса листа или хвои за 1 ч)
Породы | Освещенность, % от полного солнечного |
||
Хвойные светолюбивые: | |||
сосна | 0,08 | 3,3- |
|
лиственница | 0,06 | ||
Хвойные теневыносливые: | |||
ель | 0,06 | ||
пихта | 0,13 | ||
Лиственные светолюбивые: | |||
дуб | 0,12 | ||
ива | 0,03 | ||
береза | 0,18 | ||
Лиственные теневыносливые: | |||
клен | 0,54 | ||
липа | 0,69 |
Примечание: Знак минус перед цифрой означает, что при этом наблюдалось не поглощение, а выделение СО2 за счет дыхания.
Более слабый фотосинтез у хвойных компенсируется более длительным периодом их ассимиляционной деятельности (неопадающая хвоя) приблизительно на 3 мес., и поэтому общая продуктивность у хвойных и лиственных оказывается примерно равной.
Познакомившись коротко с влиянием света на фотосинтез, посмотрим теперь, как связан этот процесс с содержанием в атмосфере углекислоты - основного «сырья» для построения тела дерева. Ивестно, что воздух содержит всего лишь 0,03 % углекислого газа, или 3 части на 10 тыс. частей воздуха. В процессе эволюции растения приспособились усваивать углекислый газ даже при таком его содержании в воздухе. Лист дерева является очень эффективным органом и для поглощения углекислого газа. Углекислый газ поступает в лист через устьичные щели. Хотя площадь всех устьичных щелей составляет всего лишь 1 % площади листа, углекислый газ устремляется в них с очень большой скоростью.
Несмотря на все это, увеличение содержания СО2 в воздухе в 10 раз (с 0,03 до 0,3 %) усиливает фотосинтез хвои сосны. Это свойство используется при выращивании растений в оранжереях, теплицах, а также в открытом грунте. Углекислый газ можно подводить по трубам от близлежащих промышленных предприятий, выбрасывающих огромные количества его в воздух. При этом следует предварительно очистить промышленную углекислоту от содержащихся в ней вредных примесей (сернистого газа и др.). Успешный рост растений в парниках зависит не только от повышения температуры, но и от обильного питания углекислотой, выделяемой разлагающимся навозом. Следует помнить, что слишком высокая концентрация СО2 (1-2 % и выше) вредна не только для человека, но и для растений. Надо также иметь в виду, что максимального эффекта от подкормки углекислотой можно добиться лишь при достаточном освещении. Это особенно важно учитывать в теплицах и оранжереях, где нередко не хватает света.
В лесу почва постоянно выделяет СО2 в процессе разложения органических веществ различными микроорганизмами (дыхание почвы). Вследствие " этого в нижних слоях воздуха содержание СО2 может быть выше обычного (до 0,08 %). Это помогает выживать в таких условиях теневыносливым растениям, компенсируя до некоторой степени острый недостаток света, который они испытывают. В более высоких слоях воздуха (в зоне крон деревьев) в летние дни наблюдается снижение СО2 (до 0,02 % и менее) вследствие интенсивного поглощения его в процессе фотосинтеза. Ночью эта убыль вновь пополняется, так как фотосинтез прекращается, а идет только дыхание почвы и деревьев, сопровождающееся выделением углекислого газа в воздух.
Суточный ход фотосинтеза можно выразить в виде кривой. Начинаясь с восходом солнца, фотосинтез обычно достигает максимума в утренние или околополуденные часы, в полдень он падает вследствие перегрева листа и переполнения его крахмалом. У молодых деревьев (в стадии жердняка) фотосинтез идет наиболее интенсивно. Этот период обычно совпадает и с наилучшим ростом дерева. У стареющего дерева фотосинтез, а вместе с ним и нарастание органической массы (прирост) постепенно снижаются.
Как же связан фотосинтез с накоплением (приростом) органического вещества, а значит, и с урожаем? Поскольку фотосинтез - единственный процесс, создающий органическое вещество (более 90% сухого веса дерева), казалось бы, прирост и урожай должны находиться в прямой зависимости от фотосинтеза. На самом же деле все обстоит сложнее. Прежде всего, в дереве одновременно с фотосинтезом идет противоположный процесс - дыхание, заключающееся в расходе органического вещества. Поэтому в простейшем случае прирост и урожай представляют собою разницу между фотосинтезом и дыханием. Кроме того, сам прирост органического вещества в дереве делится на основной - прирост наиболее ценной части дерева, ствола, и прирост других частей (листьев, хвои, ветвей, почек, цветков, плодов и пр.), на создание которых тратится органическое вещество, но они не входят в хозяйственно ценную часть лесного дерева,
Интенсивность фотосинтеза в природных условиях меняется мало. Наибольшей она бывает у растений в крайних условиях существования (в пустынях, горах и т. д.). Поэтому прирост и урожай дерева создаются не за счет интенсивности фотосинтеза, а за счет его рабочей площади. На первый взгляд за эту величину надо принять поверхность всех листьев (или хвои) дерева. На самом же деле многие листья (нижние, находящиеся в глубине кроны ближе к стволу) не только не дают прибыли, но часто сами являются потребителями органических веществ, так как их ассимиляция не покрывает даже расходов на собственное дыхание. Вследствие этого рабочая поверхность фотосинтеза у дерева обычно бывает меньше листовой.
Действительное рабочее время фотосинтеза также оказывается меньше возможного, за которое иногда принимают весь световой день. В течение дня лист не всегда работает с накоплением органического вещества. В жаркие летние дни, особенно в полдень, листья часто вместо поглощения начинают выделять углекислоту (так же, как это бывает при недостатке света). Дождливое время тоже следует вычесть из общего рабочего времени, сюда же надо отнести весь период с температурой воздуха ниже -6°С.
Дыхание дерева слагается из средней интенсивности дыхания, массы дышащих частей и времени дыхания. Следует отметить, что масса дышащих частей и время дыхания во много раз превышают соответствующие величины для фотосинтеза. Ведь фотосинтез происходит только днем и только в листьях или хвое, да и то, как мы видели, не во всех. Дыхание же идет непрерывно во всех живых клетках каждой части дерева: листьях, хвое, ветвях, стволе и корнях.
Отсюда становится очевидным, что для накопления органического вещества в дереве фотосинтез должен в несколько раз превышать дыхание. Величина же затрат на дыхание значительна и составляет. 20-30 и даже 50 % общего количества органических веществ, созданных за счет фотосинтеза.
На этом основании некоторые ученые считают, что прирост и урожай органического вещества в дереве легче увеличить мерами, ограничивающими дыхание, чем мерами, стимулирующими фотосинтез. К сожалению, пока нет мер, которые смогли бы ограничить дыхание, по крайней мере в лесу. В теплицах же и оранжереях при подкормке растений углекислотой не только увеличивается интенсивность фотосинтеза, но одновременно и снижается дыхание, так как углекислый газ, являясь продуктом дыхания, задерживает этот процесс.
Самая большая экосистема.
гидросфера
атмосфера
биосфера
Биосфера – это геологическая оболочка Земли, охватывающая часть атмосферы, всю гидросферу и верхнюю часть литосферы вместе с населяющими их организмами. Биосфера – самая большая экосистема, объединяющая в единый планетарный круговорот отдельные круговороты веществ каждой из экосистем.
Жизненные среды биосферы.
водная, почвенная
наземно-воздушная среда
оба ответа правильные

В пределах биосферы можно выделить четыре основные среды обитания. Это водная,наземно-воздушная, почвенная среда и образуемая самими живыми организмами . Вода служит средой обитания для многих организмов. Из воды они получают все необходимые для жизни вещества: пищу, воду, газы. Поэтому, как бы ни были разнообразны водные организмы, все они должны быть приспособлены к главным особенностям жизни в водной среде. Эти особенности определяются физическими и химическими свойствами воды. Наземно-воздушная среда , освоенная в ходе эволюции позже водной, более сложна и разнообразна, и её населяют более высокоорганизованные живые организмы. Наиболее важным фактором в жизни обитающих здесь организмов являются свойства и состав окружающих их воздушных масс. Плотность воздуха гораздо ниже плотности воды, поэтому у наземных организмов сильно развиты опорные ткани — внутренний и наружный скелет. Формы движения очень разнообразны: бегание, прыгание, ползание, полёт и др. в воздухе летают птицы и некоторые виды насекомых. Потоки воздуха разносят семена растений, споры, микроорганизмы. Жизнь почвы необычайно богата. Некоторые организмы проводят в почве всю свою жизнь, другие — часть жизни. Условиями жизни в почве во многом определяются климатическими факторами, важнейшим из которых является температура. Тела многих организмов служат жизненной средой для других организмов. Условия жизни внутри другого организма характеризуются большим постоянством по сравнению с условиями внешней среды. У них не развиты органы чувств или органы движения, зато возникают приспособления для удерживания в теле хозяина и эффективного размножения.
Явление, при котором вещество передаётся по замкнутым циклам, многократно циркулируя между организмами и окружающей средой.
пищевая цепь
круговорот веществ
нет правильного ответа

Биосферный круговорот непременно включает живые и неживые компоненты. Органическое вещество может быть вновь использовано растениями только после разложения редуцентами до неорганических составляющих. Связь между живым и неживым веществом в биосферном круговороте осуществляет миграция химических элементов, входящих в состав как органических, так и неорганических соединений.
Основной источник энергии в биосфере.
Солнце
залежи нефти
продуценты
Основным источником энергии для поддержания жизни в биосфере является Солнце. Его энергия преобразуется в энергию органических соединений в результате фотосинтетических процессов, происходящих в фототрофных организмах. Энергия накапливается в химических связях органических соединений, служащих пищей растительноядным и плотоядным животным. Органические вещества пищи разлагаются в процессе обмена веществ и выводятся из организма. Выделенные или отмершие остатки разлагаются бактериями, грибами и некоторыми другими организмами. Образовавшиеся химические соединения и элементы вовлекаются в круговорот веществ. Биосфера нуждается в постоянном притоке внешней энергии, т.к. вся химическая энергия превращается в тепловую. Поэтому запасание растениями солнечной энергии в органических веществах играет исключительно важную роль в распределении и численности живых организмов.
Залежи нефти, каменного угля, торфа образовались в процессе круговорота:
азота, водорода
кислорода
углерода

В палеозойской эре происходит начальный этап накапливания нефти и газа органического происхождения углерода . В каменноугольном периоде на суше широко распространились леса, состоящие главным образом из папоротников и хвощей. Именно из упавших в воду стволов деревьев, не подвергающихся гниению, образуются огромные запасы каменных углей.
Бактерии, расщепляющие мочевину до ионов аммония и углекислого газа, принимают участие в круговороте...
азота и углерода
фосфора и серы
кислорода и углерода

Одной из специальных групп аммонификаторов являются бактерии, разлагающие мочевину. Мочевина — главная составная часть мочи человека и большинства животных. Человек выделяет бактерии, разлагающие в день от 30 до 50 г мочевины. Под влиянием бактерий мочевина разлагается, образуется карбонат аммония. Последний быстро распадается на воду, аммиак и углекислый газ .
В основе круговорота веществ лежат такие процессы, как...
расселение видов
фотосинтез и дыхание
естественный отбор

Естественным источником углерода, используемого растениями для синтеза органического вещества, служит углекислота, входящая в состав атмосферы или находящаяся в растворённом состоянии в воде. В процессе фотосинтеза углекислота превращается в органическое вещество, служащее пищей животным. Дыхание , брожение и сгорание топлива возвращают углекислоту в атмосферу.
Клубеньковые бактерии включают в круговорот...
углерод
фосфор
азот

Циркуляция биогенных элементов обычно сопровождается их химическими превращениями. Нитратный азот , может превращаться в белковый, затем переходить в мочевину, превращаться в аммиак и вновь синтезироваться в нитратную форму под влиянием микроорганизмов. В биохимическом цикле азота действуют различные механизмы, как биологические, так и химические.
Солнечная энергия улавливается...
продуцентами
редуцентами
консументами первого порядка

Лишь зелёные растения способны фиксировать световую энергию и использовать в питании простые неорганические вещества. Такие организмы выделяют в самостоятельную группу и называют автотрофами , или продуцентами — производителями биологического вещества. Они являются важнейшей частью любого сообщества, потому, что практически все остальные организмы прямо или косвенно зависят от снабжения веществом и энергией, запасёнными растениями. На суше автотрофы — это обычно крупные растения с корнями, тогда как в водоёмах их роль берут на себя микроскопические водоросли, обитающие в толще воды (фитопланктон).
Усилению парникового эффекта, по мнению учёных, в наибольшей степени способствует:
озон
углекислый газ
двуокись азота

Озон, который образует озоновый экран, формируется в:
гидросфере
мантии Земли
атмосфере

Первые живые организмы развивались в воде, которая защищала их от воздействия ультрафиолетовых лучей. Кислород, выделявшийся в процессе фотосинтеза, в верхних слоях атмосферы под действием ультрафиолетовых лучей превращался в озон (его молекула содержит три атома кислорода — О 3). По мере накопления озона произошло образование озонового слоя, который как экран, надёжно защитил поверхность Земли от губительных для живых организмов ультрафиолетовой солнечной радиации. Это позволило живым организмам выйти на сушу и заселить её.
Наибольшее количество видов находится в экосистемах:
влажного тропического леса
тайги
листопадных лесов умеренного пояса

В наши дни на Земле известно около 500 тыс. видов растений, и каждый год ботаники открывают новые. Разнообразие видов растений (флористическое) существенно различается в природных регионах планеты. Очевидно, что в пустынях видов гораздо меньше, чем в джунглях. Но как определить, где больше видов — в степях или в лесах и почему, например, в вечнозелёных тропических лесах их больше, чем в широколиственных. На эти вопросы отвечает наука биогеография, которая изучает географические закономерности формирования биологического разнообразия на Земле. Для того чтобы оценить, какие территории бедны видами, а какие богаты, составляют карты биоразнообразия. На них разными цветами отображают области с различным числом видов, приходящихся на единицу площади.
Конкретной (или локальной) флорой называют количество высших сосудистых растений на площади примерно в 100 км 2 . На островах Франца-Иосифа в приполярной области оно не превышает 50-100 видов, в тундре составляет 200-300, в тайге — 400-600, в лесостепи достигает 900 видов, в степях — 900-1000, в тропиках — более 1000.
Наиболее опасной причиной обеднения биологического разнообразия — важнейшего фактора устойчивости биосферы — является...
химическое загрязнение среды
прямое истребление
разрушение мест обитания

Одна из особенностей живого вещества.
способность быстро занимать всё свободное пространство
способность к размножению
способность к фотосинтезу
К основным особенностям живого вещества относится:
- Способность быстро осваивать все свободное пространство.
- Движение не только пассивное, но и активное.
- Устойчивость при жизни и быстрое разложение после смерти.
- Высокая адаптация к различным условиям.
- Высокая скорость протекания реакций.